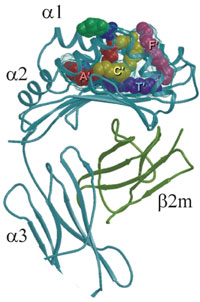
The human CD1b groove is composed of four pockets – A', C', F', and T'. Here the two alkyl chains (C16 and C18) of phosphotidylinositol lie in the A' (red) and C' (yellow) pockets, respectively. Two C16 molecules lie in the T' tunnel (violet) and the inositol ring (dark green) protudes from the groove and lies on the predicted region for T-cell receptor binding. (From D.B. Moody, S.A. Procelli,
The discovery that lipids can serve as antigens first stunned the immunological community a decade ago. But following the initial shock that floppy, hydrophobic hydrocarbon chains could stir a T-lymphocyte reaction with the same specificity as the peptides presented by conventional major histocompatibility complex (MHC) molecules, researchers hunkered down to determine the new mechanisms.
Now the process and the cast of players, including presentation molecules, loading molecules, and previously undescribed populations of...
PRECEDENTED SPECIFICITY
Specificity begins with localization. CD1 molecules serve as primary presentation modules for lipids. At least four different CD1 structures (a-d) survey endosomal compartments for lipids. And, says Mitchell Kronenberg, president and scientific director of the La Jolla Institute of Allergy and Immunology, "each of the four human CD1 molecules traffics a little bit differently inside the cell." CD1b, for example, goes into very late endosomes or even lysosomes, while CD1a is predominantly found in early endosomes. This makes sense, says Kronenberg. "There's probably some advantage for being able to survey all these different intracellular vesicles for weird glycolipids."
The CD1 molecule itself resembles MHC class I and class II molecules. The domain structure consists of a heavy chain that pairs with β-2 microglobulin. "It's very similar in 3D conformation," says Kronenberg, "except its groove for binding antigen is narrower, deeper, and very hydrophobic," great, he says, binding lipids.
But lipids were never considered antigen contenders until Michael Brenner and colleagues at Brigham and Women's demonstrated that CD1-restricted T cells were recognizing mycolic acid, the most abundant glycolipid in the cell wall of
Crystal structures of CD1 molecules reveal a few clues about binding specificity. Both length and number of lipid tails dictate how the antigen will be handled, says Brenner. Take ceramides, which have two tails: one with an 18-carbon chain, and the other with 24 carbons (with some variation). "In CD1d molecules, there are two hydrophobic pockets or channels," he says. "And the 18–20 C just fits like a ruler, just exactly. Lipid tails that are too long won't fit, and lipid tails that are too short might not anchor well."
One of mycolic acid's two tails runs more than 50 carbons long, seemingly too long to fit. But CD1b has four channels, and they're interconnected, says Brenner. "The long chain snakes sequentially through several of these interconnected channels, and sits with room left to spare," he says.
But how such molecules make it from being safely nestled within a cellular membrane to an immune presentation module is another problem. To traverse the extra-cellular fluid, lipids are carried by transfer proteins called saposins, which then participate in loading them onto CD1 molecules.2 "What we think is that these lipid-transfer proteins can essentially bind with lipids sitting in the membrane and pluck them out," says Luc Van Kaer, professor of microbiology and immunology at Vander-bilt University in Nashville, Tenn.
A HOST DEFENDER
In bacterial infection, the CD1 pathway complements the conventional MHC pathway. Both bacterial peptides and lipids are recognized by subpopulations of T cells. "We know the glycolipid antigens that are recognized by T cells in mycobacterial infections, notably TB [tuberculosis]," says Stefan Kaufmann, director of immunology at the Max Planck Institute for Infection Biology in Berlin.
© 2003 Macmillan Magazines Ltd.
CD8+ and CD4+ T cells express diverse T-cell receptors (TCRs) to recognize peptide antigens presented by MHC class I and class II molecules, respectively. By contrast, natural killer T (NKT) cells express a semi-invariant TCR that recognizes glycolipid antigens presented by CD1d molecules. (From L. Van Kaer,

Presently, 2 billion people are infected with the intracellular bacterium
CD1-restricted T cells compose between 2% and 5% of the total activated T-cell population. That may sound trivial, but says Kaufmann, "I believe that a robust beast like TB needs everything mobilized that may play a role." A 5% contribution by CD1-restricted T cells may be biologically significant against "the disease that kills more adults than any other infectious disease."
While few T cells respond to lipids, many researchers think they may have a disproportionate influence because of a quicker response time. The speed may result in part because "immature dendritic cells are capable of processing lipids onto CD1a, b, and c," says Van Kaer. "For [MHC] class II, you need some maturation going on."
BETWEEN INNATE AND ACQUIRED
The speed with which lipid antigens elicit a response also has to do with the unique set of effector cells that can be activated. The CD1d molecule, found in humans and mice, activates T cells that contain a typical T-cell receptor, but also share properties of natural killer (NK) cells. Known as NKT cells, they are activated "almost immediately," notes Brenner. In contrast, typical T cells may take a week to become effector cells. Timewise, "the NKT cell fits more in the innate category," says Brenner.
The other NKT aspect that blurs the distinction between innate and acquired immunity, says Kaufmann, is "their highly conserved T-cell receptor." While NKT cells' receptors, like those that recognize MHC-presented antigens, are generated by somatic DNA-rearrangement, they are semi-invariant and likely recognize a narrow range of antigenic diversity, more akin to Toll-like receptors of the innate immune system. "Here you have a cell that uses a highly complicated recombination receptor as something like an innate receptor," Kaufmann adds.
The first antigen identified for NKT cells didn't come from bacteria, but rather from marine sponges. While screening natural substances with antitumor properties, Japanese researchers reported that α-galactosylceramide (α-GalCer) prevents metastases in mice because it stimulated NKT cells directly.3 Since then, three clinical trials of the compound have been done with "moderate success," says Kronenberg. "Humans have the same specificity as mice," he adds, which is very unusual because typically antigen-presenting molecules are more variable. "In the case of CD1d and glycolipids, it's completely conserved. So everyone thinks it must be really important."
In addition to antitumor activity, many groups study α-GalCer against a variety of autoimmune diseases. "Generally speaking, repeated injection of α-GalCer into mice appears to promote T helper 2 responses," says Van Kaer. "Consistent with that α-GalCer does prevent type I diabetes and multiple sclerosis and rheumatoid arthritis in most of the mouse models that have been studied."
While α-GalCer has been a useful research tool and continues to be tested as a therapeutic, the search for natural antigens continues. "Until about a year ago, all we knew about NKT cells is that they see marine sponges," says Kronenberg. "No one could believe that we have conserved populations of T cell to protect us from marine sponges."
In the past year, two groups have identified physiologic antigens, presented by CD1d and activating NKT cells. One endogenous antigen is isoglobotrihexosylceramide (iGb3), a membrane glycolipid that resides in late endosomes and lysosomes.4 "So it's in the right place in the cell," says Kronenberg. But "no one knows yet how the expression of this might be regulated or why this might be important."
A bacterial antigen comes from
Although no one had predicted that lipids could serve as antigens, says Brenner, "it never made sense" that of all the potential antigens present in microbes, that only proteinaceous ones should be recognized by T cells. Now lipid antigens present a major challenge to the distinctions between innate and acquired immunity. The next surprise immunity may serve up is anyone's guess.
Interested in reading more?




