
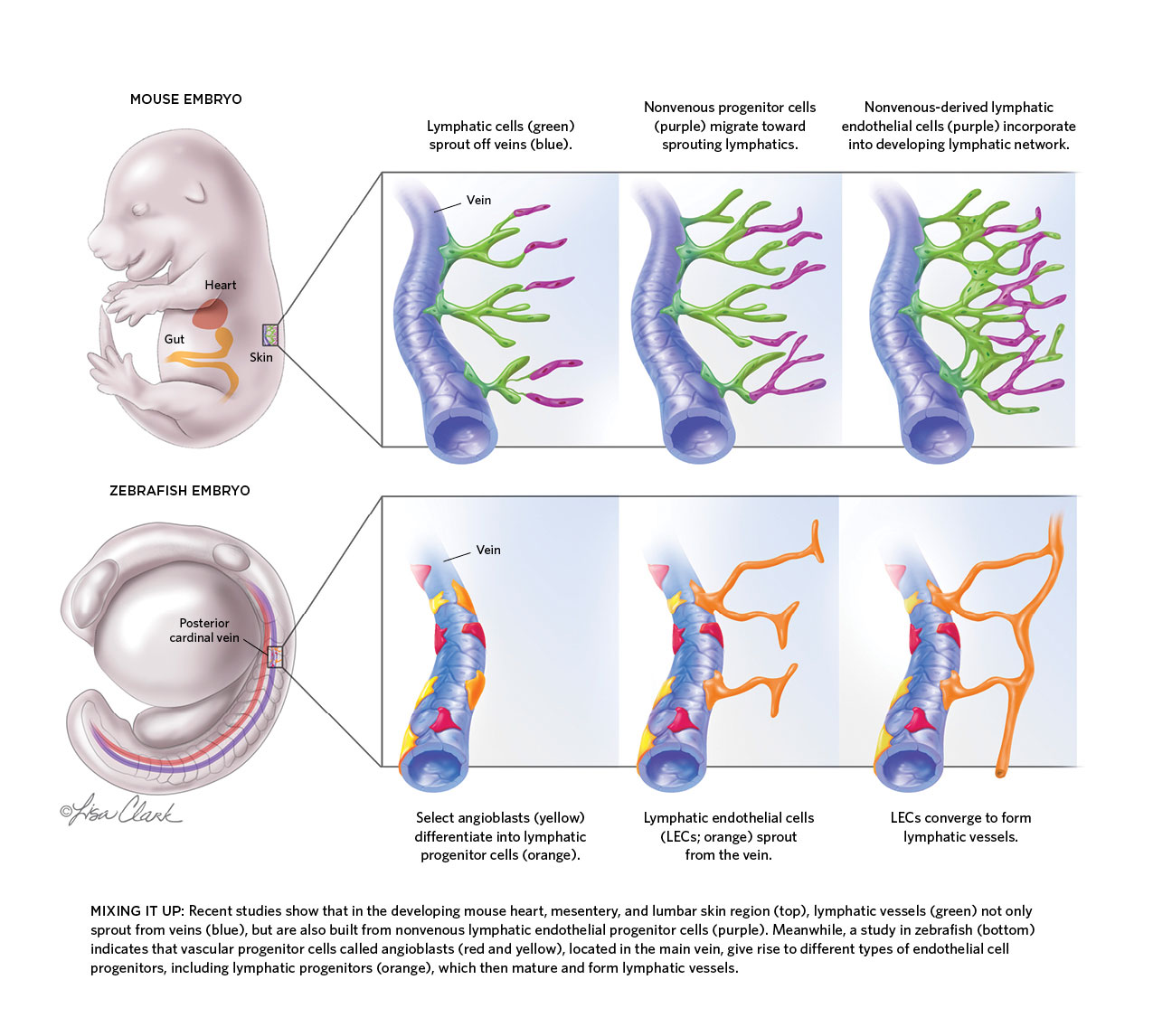
 MIXING IT UP: Recent studies show that in the developing mouse heart, mesentery, and lumbar skin region (top), lymphatic vessels (green) not only sprout from veins (blue), but are also built from nonvenous lymphatic endothelial progenitor cells (purple). Meanwhile, a study in zebrafish (bottom) indicates that vascular progenitor cells called angioblasts (red and yellow), located in the main vein, give rise to different types of endothelial cell progenitors, including lymphatic progenitors (orange), which then mature and form lymphatic vessels.
MIXING IT UP: Recent studies show that in the developing mouse heart, mesentery, and lumbar skin region (top), lymphatic vessels (green) not only sprout from veins (blue), but are also built from nonvenous lymphatic endothelial progenitor cells (purple). Meanwhile, a study in zebrafish (bottom) indicates that vascular progenitor cells called angioblasts (red and yellow), located in the main vein, give rise to different types of endothelial cell progenitors, including lymphatic progenitors (orange), which then mature and form lymphatic vessels.
See full infographic: JPG© LISA CLARK
For 10 years, Karina Yaniv has worked to find out just how much zebrafish have in common with mice—at least when it comes to their lymphatic systems, the open-ended networks of vessels best known for draining fluids from tissues and providing thoroughfares for immune cells throughout the body. Yet in doing so she ended up discovering something that had very little in common with the findings of numerous earlier studies on other animals’ lymphatic systems. Contrary to the widely held view in developmental biology, she found, lymphatics don’t always originate from veins.
“It was scary—I’m a young PI, so it’s not easy to stand up against the stream,” says Yaniv, who studies developmental biology at the Weizmann Institute of Science in Rehovot, Israel. But it ...














{kind=link}