
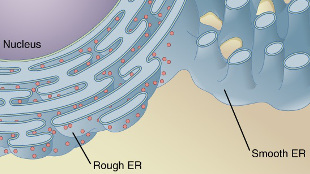
 Illustration from Anatomy & PhysiologyWIKIMEDIA, OPENSTAX COLLEGEThe endoplasmic reticulum (ER) has specific receptors that target it for autophagy, or intracellular recycling, according to two independent studies published this month (June 3) in Nature. Hitoshi Nakatogawa of the Tokyo Institute of Technology led a team that identified these receptors in yeast. Ivan Dikic of the Goethe University School of Medicine led a team that examined the same kinds of receptors in mice.
Illustration from Anatomy & PhysiologyWIKIMEDIA, OPENSTAX COLLEGEThe endoplasmic reticulum (ER) has specific receptors that target it for autophagy, or intracellular recycling, according to two independent studies published this month (June 3) in Nature. Hitoshi Nakatogawa of the Tokyo Institute of Technology led a team that identified these receptors in yeast. Ivan Dikic of the Goethe University School of Medicine led a team that examined the same kinds of receptors in mice.
“The major finding is that ER . . . has a homeostatic mechanism to remodel itself, to turn over its own contents,” said Dikic. “And that this mechanism, which regulates this housekeeping function, is mediated by selective autophagy.”
“Both studies report a mechanism that cells can use to selectively sequester the ER for degradation via autophagy. This is accomplished by receptors that the studies identify that bind to the ER and also to components of the autophagosome,” David Rubinsztein of the Cambridge Institute for Medical Research, who penned a commentary accompanying the studies but was not involved in the work, wrote in an e-mail to The Scientist.
Autophagy is a process by which eukaryotic cells break down and recycle their own materials. In autophagy, a ...













{kind=link}